尋媶撪梕
B嵶朎弶婜暘壔偺惂屼婡峔
丂B嵶朎偼憿寣姴嵶朎偐傜敪惗偟丄僾儘B嵶朎丄僾儗B嵶朎傪宱偰丄B嵶朎傊偲暘壔偟傑偡丅僸僩傗儅僂僗側偳懡偔偺摦暔偱偼丄戀帣婜偵偼庡偵娞憻偱丄惗屻偼庡偵崪悜偱偙偺B嵶朎弶婜暘壔偑婲偙傝傑偡丅B嵶朎偺弶婜暘壔偱嵟傕摿挜揑側偙偲偼峈懱乮柶塽僌儘僽儕儞丗Ig乯堚揱巕偺嵞峔惉偑婲偙傞偙偲偱偡丅傑偢丄H嵔堚揱巕椞堟偵偁傞懡悢偺V, D, J偺堚揱巕抐曅偐傜偦傟偧傟侾偮偢偮偑儔儞僟儉偵慖偽傟偰DNA慻傒姺偊偵傛傝寢崌偟丄VDJ偲偄偆傂偲偮偺僄僋僜儞傪宍惉偟傑偡丅偙偺夁掱偼僾儘B嵶朎偱婲偙傝丄偙偺嵞峔惉偵惉岟偟偨H嵔堚揱巕偐傜偼兪H嵔抈敀偑嶌傜傟傑偡丅兪H嵔抈敀偼偦傟埲慜偵敪尰偟偰偄偨VpreB偍傛傃兩5偲偄偆傆偨偮偺抈敀偲偲傕偵暋崌懱乮僾儗B嵶朎庴梕懱丗pre-B cell receptor, preBCR乯傪宍惉偟傑偡丅VpreB偲兩5偼偁傢偣偰L嵔條偺峔憿傪傕偭偰偄傞偺偱戙懼L嵔乮surrogate light chain乯偲傕屇偽傟傑偡偑丄B嵶朎埲崀偺暘壔抜奒偱偼敪尰偟偰偄傑偣傫丅preBCR偑嵶朎枌忋偵敪尰偡傞偲嵶朎暘楐偑婲偙傝丄RAG妶惈偺掅壓偵傛傝H嵔嵞峔惉偑巭傑傝丄嵶朎悢偑憹偊傑偡丅偙偺抜奒偺嵶朎傪戝宆僾儗B嵶朎偲屇傃傑偡丅幚嵺偵偼傎偲傫偳偺戝宆僾儗B嵶朎偼preBCR堿惈偱偡丅偙傟偼preBCR偑嵶朎昞柺偵敪尰偟偰偄傞帪娫偑旕忢偵抁偄偨傔偲峫偊傜傟偰偄傑偡丅悢夞偺嵶朎暘楐偺屻丄暘楐偼掆巭偟丄嵶朎偼彫宆僾儗B嵶朎偲側傝傑偡丅偙偺嵶朎偱偼L嵔乮內 / 兩乯堚揱巕偺V-J嵞峔惉偑婲偙傝傑偡丅偙傟偵惉岟偟偨堚揱巕偐傜嶌傜傟傞L嵔抈敀偼兪H嵔偲堦弿偵IgM傪宍惉偟偰丄嵶朎枌忋偵敪尰偟傑偡丅偙偙偱偮偄偵IgM偲偄偆峈尨庴梕懱傪帩偭偨B嵶朎偑抋惗偟傑偡*丅師偵B嵶朎偼偙偺IgM偲摨偠峈尨摿堎惈傪帩偮IgD傪傕敪尰偟丄崪悜偐傜枛徑儕儞僷慻怐傊偲堏峴偟傑偡丅枛徑偱偼偝傜偵丄IgM傛傝IgD偺敪尰偑崅偄惉弉B嵶朎傊偲暘壔偟傑偡丅
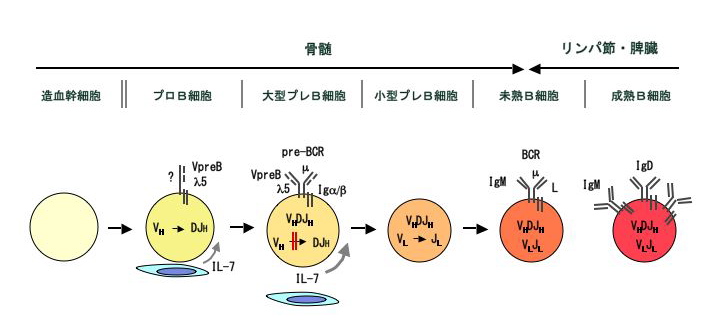
丂preBCR傪嵶朎昞柺偵敪尰偟偰偄傞嵶朎偼旕忢偵彮側偄偺偱偡偑丄枌宆兪嵔曄堎儅僂僗偱偼B嵶朎暘壔偑僾儗B嵶朎偺庤慜偱掆巭偟偰偄傞偺偱丄傗偼傝嵶朎枌忋偵preBCR偑敪尰偟偰嵶朎撪偵僔僌僫儖傪憲傞偙偲偑廳梫偩偲峫偊傜傟傑偡丅preBCR傪峔惉偡傞枌宆兪嵔丄兩5丄VpreB偺偦傟偧傟偺僲僢僋傾僂僩儅僂僗偺夝愅偐傜丄preBCR偵偼埲壓偺傛偆側婡擻偑偁傞偙偲偑柧傜偐偵側偭偰偄傑偡丅
丂丂侾乯 IgH嵔堚揱巕嵞峔惉偺梷惂乮懳棫堚揱巕攔彍乯
丂丂俀乯僾儗B嵶朎偺暘楐丒憹怋
丂丂俁乯嵶朎昞柺偺preBCR偺徚幐
丂丂係乯戝宆僾儗B嵶朎偐傜彫宆僾儗B嵶朎傊偺暘壔
丂丂俆乯L嵔堚揱巕嵞峔惉偺懀恑
丂偙傟傜偺婡擻偺惗棟揑側堄枴偵偮偄偰埲壓偵愢柧偟傑偡丅
侾乯 堦曽偺H嵔懳棫堚揱巕乮傾儗儖乯偑V-DJ寢崌偵惉岟偟婡擻揑側兪H嵔傪敪尰偡傞偲傕偆堦曽偺傾儗儖偺嵞峔惉傪梷惂偡傞偲偄偆傕偺偱丄B嵶朎偺乽侾嵶朎乗侾摿堎惈乿偺尨懃傪妋棫偡傞偺偵廳梫側婡峔偱偡丅偄傑丄DJ寢崌傑偱恑傫偩H嵔堚揱巕偺俀偮偺傾儗儖偑帺桼偵V-DJ嵞峔惉偱偒傞偲偡傞偲丄V偲DJ偺僐僪儞偺撉傒榞偑堦抳偡傞妋棪偼侾/俁偩偐傜丄偛偔扨弮偵峫偊偰侾/俋偺妋棪偱椉曽偺傾儗儖偑婡擻揑側H嵔堚揱巕傪嶌偭偰偟傑偄傑偡丅堦曽丄椉曽偺傾儗儖偑偲傕偵旕婡擻揑側H嵔堚揱巕傪嶌傞妋棪偼俀/俁亊俀/俁=係/俋偱丄偙偺傛偆側嵶朎偼峈尨庴梕懱傪敪尰偟側偄偺偱B嵶朎偵偼側傟傑偣傫丅偟偨偑偭偰丄嶻惗偝傟傞B嵶朎偺偆偪俀偮偺婡擻揑側H嵔堚揱巕傪傕偮嵶朎偼侾/俋亐俆/俋=侾/俆偲側傝傑偡丅偡側傢偪丄懳棫堚揱巕攔彍偺婡峔偑側偗傟偽丄俀侽亾偺B嵶朎偺偦傟偧傟偑俀偮偺堎側傞H嵔傪敪尰偟偰偟傑偄丄俀偮偺峈尨摿堎惈傪帩偮偙偲偵側傝傑偡丅偙傟傜偺嵶朎偑偁傞峈尨偵弌夛偭偰偦傟偵懳偡傞峈懱傪嶻惗偡傞傛偆偵側傞偲丄傕偆傂偲偮偺柍娭學偺摿堎惈傪帩偮峈懱傕嶻惗偝傟偰丄柶塽墳摎偺摿堎惈偑幐傢傟偰偟傑偄傑偡丅
俀乯 婡擻揑側H嵔抈敀偑偱偒偨偙偲傪僠僃僢僋偟丄偦偺婡擻揑側嵶朎傪慖戰揑偵憹壛偝偣傞偺偵栶棫偭偰偄傑偡丅DNA慻傒姺偊偵傛傞Ig堚揱巕嵞峔惉偼惉岟傛傝幐攕偺曽偑偼傞偐偵懡偄僾儘僙僗偱偡偐傜丄尷傜傟偨僗儁乕僗偺拞偱嵞峔惉偵惉岟偟偨桳堄媊側嵶朎傪慖戰偡傞偙偲偼廳梫偱偡丅
俁乯 preBCR偼兩5偲Vpre儬傪夘偟偰帺屓壦嫶偡傞偙偲偵傛傝嵶朎撪偵僔僌僫儖傪憲傞偲偲傕偵丄僄儞僪僒僀僩乕僔僗偵傛偭偰嵶朎撪偵庢傝崬傑傟傑偡丅偦偺屻偼兩5,Vpre儬偺堚揱巕敪尰偑柍偔側傞偙偲偵傛偭偰preBCR偺嵞敪尰偑梷巭偝傟傑偡丅preBCR偺徚幐偼僾儗B嵶朎偺憹怋掆巭傪傕偨傜偡偲峫偊傜傟偰偄傑偟偨偑丄嵟嬤丄preBCR 傪帩懕揑偵敪尰偝偣偨僩儔儞僗僕僃僯僢僋儅僂僗偱傕僾儗B嵶朎偺憹怋丒暘壔偼惓忢偱偁傞偙偲偑帵偝傟傑偟偨丅堦曽丄偙偺儅僂僗偱偼B嵶朎偑帺敪揑偵妶惈壔偟峈懱傪嶻惗偡傞偙偲偐傜丄preBCR 偐傜偺峆忢揑僔僌僫儖偼B嵶朎偺惓忢側惂屼傪朩偘傞偲峫偊傜傟傑偡丅
係乯 暘壔偑恑峴偡傞偙偲偑廳梫側偺偼偄偆傑偱傕偁傝傑偣傫偑丄偙偙偱偼暘楐偑掆巭偟偰彫宆僾儗B嵶朎傊暘壔偟丄RAG-1/2乮Ig堚揱巕嵞峔惉偵昁恵偺峺慺乯偺妶惈偑嵞忋徃偟丄L嵔堚揱巕偺嵞峔惉偑奐巒偝傟傞偙偲傕廳梫偱偡丅傕偟丄H嵔偲L嵔偺堚揱巕偑摨帪婜偵嵞峔惉偡傞偲幐攕偺妋棪偼憡忔揑偵崅偔側傞偼偢偱偡偐傜丄偡偱偵婡擻揑H嵔傪桳偡傞嵶朎偵偍偄偰L嵔堚揱巕嵞峔惉偑奐巒偝傟傞偙偲偑岠棪傪崅傔傞偺偵廳梫偩偲巚傢傟傑偡丅
俆乯 preBCR傪敪尰偱偒側偄僲僢僋傾僂僩儅僂僗偱傕L嵔堚揱巕偺嵞峔惉偼婲偙傝傑偡偑丄偦偺昿搙偼掅偄偙偲偐傜丄preBCR僔僌僫儖偼L嵔堚揱巕偺嵞峔惉傪懀恑偡傞偲峫偊傜傟傑偡丅preBCR僔僌僫儖偼RAG-1/2偺敪尰傪掅壓偝偣傞堦曽偱丄L嵔堚揱巕椞堟偺揮幨乮Germline transcript乯傪懀恑偟丄嵞峔惉偺偨傔偺慻姺偊峺慺傪庴偗擖傟傗偡偄忬懺偵偡傞偲峫偊傜傟偰偄傑偡乮Accessibility model)丅
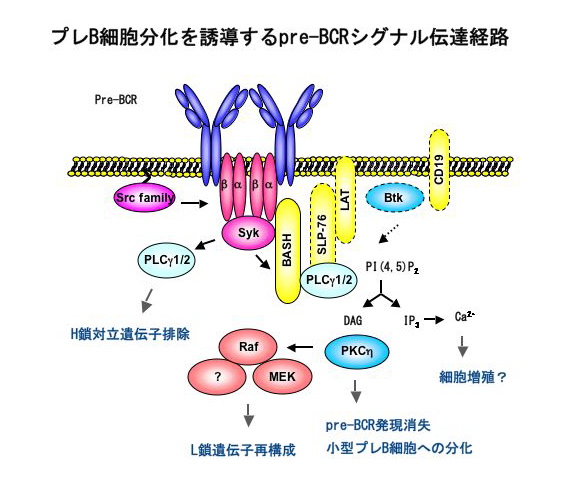
丂偙偺傛偆偵廳梫側婡擻傪扴偆preBCR偱偡偑丄偦偺僔僌僫儖揱払婡峔偼傑偩廫暘偵棟夝偝傟偰偄傑偣傫丅巹偨偪偼偙傟傜偺堎側傞婡擻偑偳偺傛偆偵偟偰堷偒婲偙偝傟傞偺偐丄偦傟偑preBCR偐傜偳偺傛偆側僔僌僫儖宱楬偱揱払偝傟傞偺偐偲偄偆偙偲傪丄暘巕儗儀儖偱柧傜偐偵偟偨偄偲峫偊偰偄傑偡丅偙傟傑偱偺尋媶偐傜丄preBCR偺僔僌僫儖揱払偵娭傢傞僔僌僫儖場巕偺傎偲傫偳偼峈尨庴梕懱乮BCR乯僔僌僫儖偺傕偺偲嫟捠偩偲峫偊傜傟偰偄傑偡丅偟偐偟丄偦傟傜偺僲僢僋傾僂僩儅僂僗偼BCR僔僌僫儖偵傛傞B嵶朎偺妶惈壔堎忢傪帵偡傕偺偑傎偲傫偳偱丄僾儗B嵶朎偺庤慜偱暘壔掆巭傪帵偟偨偺偼Syk僲僢僋傾僂僩儅僂僗偩偗偱偡丅Syk偼僔僌僫儖揱払偺婲揰偲傕偄偆傋偒僠儘僔儞僉僫乕僛偱庬乆偺僔僌僫儖場巕傪儕儞巁壔偟傑偡偑丄偦偺偆偪偳偺場巕偑偳偺宱楬偵摥偄偰偄傞偺偩傠偆偐偲偄偆偺偑巹偨偪偺媈栤偱偡丅巹偨偪偑嶌惢偟偨BLNK乮暿柤SLP-65/BASH乯偺寚懝儅僂僗偱偼B嵶朎弶婜暘壔偑晄姰慡偱丄彮悢偺B嵶朎偼懚嵼偡傞傕偺偺丄崪悜偱偼彫宆僾儗B嵶朎偑徚幐偟丄戙傢傝偵戝宆僾儗B嵶朎偑拁愊偟偰偄傑偟偨丅偍傕偟傠偄偙偲偵丄偙偺戝宆僾儗B嵶朎偼捠忢尒傜傟側偄preBCR梲惈偺嵶朎偱丄偟偐傕嵶朎夞揮偑傎偲傫偳掆巭偟偰偄傑偟偨丅偟偨偑偭偰丄暘壔桿摫偵昁梫側preBCR偺僔僌僫儖揱払偵偼BLNK偑娭傢偭偰偍傝丄preBCR傪敪尰偟偰傕BLNK偑側偄偲嵶朎暘楐偑偆傑偔桿摫偱偒偢丄偦偺抜奒偱暘壔偑掆懾偟偰偟傑偭偨偲峫偊傜傟傑偡丅傑偨丄preBCR偺嵶朎昞柺偐傜偺慺憗偄徚幐偵傕BLNK傪夘偟偨preBCR僔僌僫儖偑娭梌偟偰偄傞壜擻惈傕帵嵈偝傟傑偡丅偟偐偟側偑傜丄H嵔懳棫堚揱巕攔彍偼崪悜偱傕枛徑偱傕惓忢偱偁傝丄俀庬椶偺H嵔傪敪尰偟偰偄傞僾儗B嵶朎傗B嵶朎偼偁傝傑偣傫偱偟偨丅偙偺偙偲偐傜丄僔僌僫儖宱楬偼偡偱偵BLNK偺偲偙傠偱彮側偔偲傕俀偮偵暘婒偟偰偄傞偙偲偑柧傜偐偲側傝傑偟偨丅
丂嬃偄偨偙偲偵BLNK僲僢僋傾僂僩儅僂僗偺悢亾偵偼僾儗B嵶朎桼棃偺敀寣昦偑敪徢偟傑偟偨乮乽僾儗俛嵶朎惈媫惈儕儞僷惈敀寣昦偺敪惗婡峔乿偺崁嶲徠乯丅巹偨偪偼敪徢偟偨儅僂僗偺崪悜嵶朎傪挿婜攟梴偡傞偙偲偵傛傝偙偺敀寣昦桼棃偺僾儗B嵶朎姅傪悢姅庽棫偟傑偟偨丅偙傟傜偺嵶朎偼preBCR傪敪尰偟偰偍傝丄BLNK偺摫擖傗峈懱偵傛傞preBCR偺嫮壦嫶偵傛偭偰丄preBCR偺徚幐偲L嵔堚揱巕偺嵞峔惉丄L嵔敪尰偑桿摫偝傟傞偙偲偑傢偐傝傑偟偨丅偙偺僾儗B嵶朎姅偼偙傟傜偺僀儀儞僩傪堷偒婲偙偡preBCR僔僌僫儖揱払宱楬傪夝柧偡傞偺偵桳梡側僣乕儖偲側傞偲巹偨偪偼峫偊傑偟偨丅偦偟偰尋媶偺寢壥丄BLNK偐傜抈敀僉僫乕僛偱偁傞PKC兣傪夘偡傞僔僌僫儖偑揮幨場巕IRF-4偺敪尰傪桿摫偟偰丄偦偺寢壥丄preBCR敪尰掆巭丄L嵔堚揱巕椞堟偺揮幨乮Germline transcript乯偍傛傃嵞峔惉傪懀恑偡傞偙偲偑柧傜偐偵側傝傑偟偨丅
丂堦曽丄H嵔懳棫堚揱巕攔彍傪婲偙偡preBCR僔僌僫儖偵偼Syk傗PLC兞2偼昁梫偱偡偑丄BLNK傗Btk偼昁梫偱側偔丄PLC兞2埲壓偺僔僌僫儖宱楬偼崱偺偲偙傠暘偐偭偰偄傑偣傫丅
拲 *丂嵞峔惉偵惉岟偟偨傂偲偮偺柶塽僌儘僽儕儞H嵔堚揱巕偐傜揮幨偝傟偨RNA乮VDJ-C兪乯偐傜堎側傞僗僾儔僀僔儞僌偵傛傝俀庬椶偺mRNA偑偱偒丄偦偙偐傜枌宆兪H嵔偲暘斿宆兪H嵔偲偑東栿偝傟傑偡丅慜幰偑L嵔偲寢崌偟偨傕偺偑峈尨庴梕懱偱丄屻幰偑L嵔偲寢崌偟偨傕偺偑峈懱偱偡丅B嵶朎偑峈尨偲弌夛偆傑偱偼慜幰偺mRNA偑傎偲傫偳偱丄峈尨庴梕懱乮枌宆IgM乯偑嵶朎昞柺偵敪尰偟偰偄傑偡偑丄峈尨巋寖傪庴偗傞偲僗僾儔僀僔儞僌偺僷僞乕儞偑曄壔偟丄屻幰偺mRNA偑媫憹偟偰丄暘斿宆偺IgM峈懱偑戝検偵嶻惗偝傟傞傛偆偵側傝傑偡丅傑偨丄IgD偺H嵔偱偁傞兟嵔傕摨偠H嵔堚揱巕偐傜嶻惗偝傟傑偡偑丄兪H嵔mRNA偑VDJ偲C兪堚揱巕偲偺僗僾儔僀僔儞僌偵傛傞偺偵懳偟丄兟嵔mRNA偼VDJ偲C兟堚揱巕偲偺僗僾儔僀僔儞僌偵傛傝嶌傜傟傑偡丅
偍壣側傜偙偪傜傕 仺 乽峈尨傪擣幆偡傞丄屘偵B嵶朎嵼傝乿
Back