
研究内容
レタスの酸性条件下での根毛形成誘導機構を解明する
工場からの排気や私たちの生活から出た硫酸や窒素化合物(NOx)は大気中の水に溶け、酸性雨として降ってきます。そのpHは4付近まで低下しています。酸性雨は陸上や海洋、湖沼の生物にいくつかの異なる仕組みで影響を与えていますが、私たちの研究室では特に「低pH」に注目して研究を進めています。
下の写真左側はpH5.5で培養したレタス、右側はpH4.0で培養したレタスです。pH5.5で育てたレタスの主根は長く伸びていますが、pH4.0で育てたレタスの根は短くなっています。低pHによって主根の伸長が阻害されていることから、水の吸収能力が低下し、地上部の生長も悪くなると考えられますが、実際に写真を見てみるとほとんどどちらもほとんど大きさは変わりません。このことから私たちは、レタスには酸性条件に対応する何らかの仕組みが備わっていると考えました。光学顕微鏡や電子顕微鏡を用いた実験などにより、酸性条件下では主根が短くなった代わりに、表皮細胞に大量の根毛を形成し、表面積を大きくすることで水の吸収能力を補っていることがわかりました。
現在は酸性条件下での根毛形成誘導機構について、生理学的手法や、分子生物学的手法を用いて解析を行っています。
根毛形成と植物ホルモン
植物が根毛を形成するのには根の表面積を大きくして、水分や各種必要な無機塩をより吸収しやすくしているといわれています。根毛形成時には植物ホルモンのオーキシンやエチレンが促進的にはたらいていることが主にシロイヌナズナの突然変異体を用いた実験などによる研究で明らかにされています。私たちの研究室では、主にレタスにおいて無機塩培地で水耕栽培することで根毛形成と植物ホルモンの関わりについて研究しています。
pH6の培地にオーキシンや植物体内でエチレンに変換される物質を添加すると根毛形成が誘導され、pH4の条件下でオーキシンの作用を阻害したり、エチレンの生合成や作用を阻害すると根毛形成も抑制されます。また、pH4の条件下ではpH6のときとくらべてエチレンの生合成量が増加します。
以上のような実験を行い、レタスの低pHに応答した根毛形成誘導にはエチレン・オーキシンの作用が必要であることを明らかにしました。
 レタス主根の電子顕微鏡写真
レタス主根の電子顕微鏡写真
A: pH6で育てたレタス主根の表面
B: pH4で育てたレタス主根の表面
C: pH6の培地にACCを添加して育てたレタス主根の表面
1日pH6培地で育てた後pH4の培地にうつし、さらにその後9時間経過したときの主根の様子です。
主根の表皮細胞の境目が見えるよう、表面を液体窒素で一度凍らせてから低真空走査型電子顕微鏡で観察しました。
図の右下の白線は500μmを表しています。
写真をクリックするとさらに鮮明な画像を見ることができます。
さらに私たちの研究室では、エチレンの生合成に関わる遺伝子(Ls-ACS1、Ls-ACS2)を単離し、それらの発現が低pH条件によってどのように制御されているかも調べました。その結果、発現のパターンに違いはあるもののどちらも低pH条件によって発現が誘導されていることが明らかになりました。

根毛形成と細胞骨格
根毛形成には表皮細胞の表層微小管(CMT)の配向のランダム化が必須です。中性条件で細胞長軸に対して垂直に配向しているCMTは、pH低下処理後速やかにランダム化することがわかりました。さらに本研究では、このランダム化にオーキシンとエチレンがどのように関与しているかを調べました。
その結果エチレンやオーキシン、それらの阻害剤の添加実験の結果、1.低pHによるCMTランダム化にはオーキシンが中心的な役割を果たしていること、2.低pHによって誘導されるエチレンはオーキシンの作用を増幅している可能性があることなどが示されました。過去の研究からpH低下処理でエチレン生合成量は増加するが根全体のオーキシンの量は増加しないことがわかっています。
以上のことから低pHで誘導される多量のエチレンが根毛形成細胞のオーキシンに対する感受性を増加させている可能性、もしくはオーキシンの局在を変化している可能性が考えられる結果となりました。
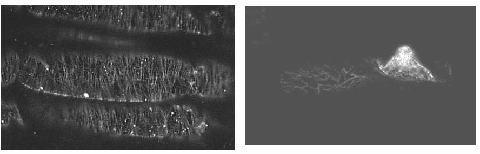
左:pH6で培養したときの表層微小管の配向
右:pH4で培養したときの表層微小管の配向
白く繊維状に見えるのが表層微小管です。pH4の写真で特に微小管が集中している部分がありますが、これは根毛原基といい、この突起の先端部分が伸びてやがて成熟した根毛になります。