峈尨傪擣幆偡傞丄屘偵B嵶朎嵼傝
丂乽変巚偆丄屘偵変嵼傝乿偲偄偆捠傝丄恖娫偺傾僀僨儞僥傿僥傿乕偼擼偑巚峫偡傞偙偲偵傛偭偰惗偠丄偦偺巚峫僷僞乕儞偑恖娫偺屄惈偱偁傞偲偄偊傞丅堦曽丄儕儞僷媴偺傾僀僨儞僥傿僥傿乕傪寛傔偰偄傞傕偺偼峈尨庴梕懱偱偁傞丅傾僀僨儞僥傿僥傿乕偑柧妋偱側偄恖娫偼懡偄偑丄傾僀僨儞僥傿僥傿乕傪妋棫偱偒側偄儕儞僷媴偼偦偺懚嵼傪嫋偝傟側偄丅偡側傢偪峈尨庴梕懱傪傑偲傕偵敪尰偱偒側偄儕儞僷媴偼偡傋偰敪惗偺夁掱偱暘壔偺恑峴偑巭傑傝丄巰偺塣柦傪扝傞丅B嵶朎宯楍偱偼丄兪H嵔偲VpreB, 兩5偐傜惉傞僾儗B嵶朎庴梕懱乮preBCR乯偑枌抈敀偲偟偰敪尰偡傞偙偲偵傛偭偰傑偢峈尨庴梕懱H嵔偺敪尰偑僥僗僩偝傟傞栿偱偁傞丅崱偱偼柶塽妛偺忢幆偲側偭偨偑丄偙傟傪弶傔偰柧妋偵帵偟偨偺偼Klaus Rajewsky偺尋媶幒偱巹偨偪偑嶌惢偟偨兪H嵔堚揱巕枌僄僋僜儞偍傛傃兩5堚揱巕偺曄堎儅僂僗偺昞尰宆偱偁偭偨丅椉幰偺崪悜偱偼丄彫宆僾儗B嵶朎傛傝慜偺抜奒偱暘壔偑掆巭偟偰偄偨丅偦傟埲慜偵偼丄僾儗B嵶朎姅偺攟梴宯偱枌宆兪H嵔偑偛偔掅昿搙偵L嵔堚揱巕偺嵞峔惉傪桿摫偡傞偙偲偑帵偝傟偰偄偨偩偗偱丄僾儗B嵶朎偵兪H嵔偺傒偑敪尰偟偰偄傞偙偲偺堄枴偼柧傜偐偱側偐偭偨丅偟偐偟丄僾儗B嵶朎摿堎揑偵敪尰偡傞L嵔條堚揱巕兩5丄VpreB偑摨掕偝傟丄偦傟傜偺嶻暔偑兪H嵔偲寢崌偟偆傞偙偲傕傢偐傝丄偙傟傜偺敪尒偑阾柧婜偺僕乕儞僞乕僎僥傿儞僌媄弍偲尒帠偵梈崌偟偰儕儞僷媴弶婜暘壔偺僷儔僟僀儉偲偟偰幚傪寢傫偩偲偄偆傢偗偩丅T嵶朎弶婜暘壔傕摨條偵b嵔傪娷傓preTCR偵傛偭偰惂屼偝傟傞偙偲偑娫傕側偔柧傜偐偵偝傟偨丅
丂preBCR偺嵶朎枌忋偱偺敪尰偼偄偔偮偐偺嵶朎姅偱偼柧傜偐偩偑丄崪悜撪偺僾儗B嵶朎偱偼枹偩偵偼偭偒傝偟側偄丅僾儗B嵶朎偺悢偑彮側偄忋偵preBCR偺敪尰検傕彮側偔丄傑偨堦夁惈偱偁傞偙偲偑専弌傪崲擄偵偟偰偄傞偲巚傢傟傞丅惓忢僾儗B嵶朎忋偱偺preBCR偺敪尰傪媈栤帇偡傞惡傕偁偭偨偑丄BCR僔僌僫儖揱払偺拞幉偱偁傞Ig兛/兝傗Syk僉僫乕僛偺曄堎儅僂僗傕preBCR寚娮儅僂僗偲摨條偺昞尰宆傪帵偟偨偙偲偐傜丄BCR偲摨條偵preBCR傕嵶朎枌忋偐傜僔僌僫儖傪敪偟偰暘壔傪桿摫偟偰偄傞偲峫偊傜傟傞乮埲慜偺曬崘偲堎側傝丄Ig兝曄堎儅僂僗偱傕VDJ嵞峔惉傑偱暘壔偼恑峴偟丄傑偨丄Ig兛 null曄堎儅僂僗偱傕兪H嵔曄堎儅僂僗偲摨偠抜奒偱暘壔偑掆巭偡傞偦偆偱偁傞丟巹怣乯丅preBCR偁傞偄偼preTCR偺寚娮偵傛偭偰掆巭偟偨B丒T嵶朎弶婜暘壔偑Ig兝傗CD3偺壦嫶偵傛傝夞暅偡傞偙偲傕偙傟傪嫮偔巟帩偡傞丅師偼偙偺preBCR偐傜偺僔僌僫儖偑偳偺傛偆偵揱偊傜傟偰偄傞偐偱偁傞丅preBCR偼僾儗B嵶朎偺暘楐偲偦偺屻偺彫宆僾儗B嵶朎傊偺暘壔偩偗偱側偔丄H嵔堚揱巕嵞峔惉偺梷惂乮懳棫堚揱巕攔彍乯傗L嵔堚揱巕嵞峔惉偺懀恑乮堎榑偼偁傞偑乯偵傕僔僌僫儖傪憲偭偰偄傞偲偝傟偰偄傞丅偙偺僔僌僫儖宱楬偑偳偺傛偆偵怳傝暘偗傜傟揱偊傜傟偰偄傞偺偐傪柧傜偐偵偡傞偺偑巹偨偪偺尰嵼偺尋媶栚昗偺傂偲偮偱偁傞丅庡偵B嵶朎姅傪梡偄偨尋媶偐傜丄Syk, Lyn, Btk偲偄偭偨僠儘僔儞僉僫乕僛偲偦傟傜偺婎幙偲側傞PLC兞2, Vav, PI3-kinase, Shc, HS1, 偦偺懠Grb2, Ras, Rac偲偄偭偨抈敀偑BCR僔僌僫儖揱払偺弶婜偵娭梌偡傞偲峫偊傜傟偰偄傞偑丄偦偺偆偪僲僢僋傾僂僩儅僂僗偺夝愅偐傜preBCR僔僌僫儖偵傛傞B嵶朎弶婜暘壔偵摥偄偰偄傞偙偲偑柧傜偐偵側偭偨偺偼堄奜偵傕Syk偺傒偱偁傞*丅
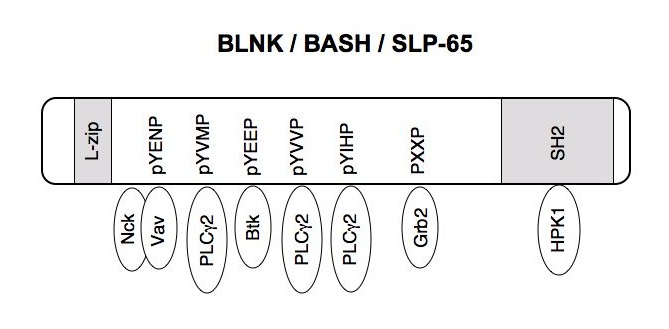
丂巹偨偪偑僯儚僩儕偺僼傽僽儕僉僂僗擷偐傜PCR-僒僽僩儔僋僔儑儞朄偵傛傝摨掕偟偨B嵶朎摿堎揑偵敪尰偡傞抈敀BASH (B-cell adaptor containing an SH2 domain) 偼丄TCR僔僌僫儖揱払偵廳梫側傾僟僾僞乕偲偟偰拲栚偝傟偰偄偨SLP-76偲傛偔帡偨峔憿傪偟偰偄偰丄幚嵺丄BCR壦嫶偵傛傝僠儘僔儞儕儞巁壔偝傟Syk偲夛崌偡傞側偳丄SLP-76偲摨條偵BCR偺壓棳偱僔僌僫儖暘巕偺傾僟僾僞乕偲偟偰婡擻偟偰偄傞偲峫偊傜傟偨丅傑偨丄BASH偺儅僂僗儂儌儘僌偲巚傢傟傞晄姰慡側cDNA傪僨乕僞儀乕僗偵尒弌偟丄偦偺慡挿傪僋儘乕儞偟偨丅偙傟偱BCR僔僌僫儖揱払尋媶傪break through偱偒傞偺偱偼偲嫽暠偟丄偡偖偵DT40嵶朎偲儅僂僗偺儈儏乕僞儞僩嶌傝傗in vitro偺幚尡偵偲傝偐偐偭偨丅偟偐偟丄嬃偔傋偒偙偲偵摨偠偙偲傪傗偭偰偄傞僌儖乕僾偑懠偵俁偮傕偁傞偙偲偑屻偵敾柧偟偨丅Andy Chan偺僌儖乕僾偼BCR巋寖屻丄PLC兞2, Vav, Grb2, Nck偲寢崌偡傞儕儞巁壔抈敀傪僸僩B嵶朎偐傜惛惢扨棧偟BLNK (B-cell linker) 偲柤晅偗偨丅傑偨丄Michael Reth偺僌儖乕僾偼BCR偺敪尰偵埶懚偟偰pervanadate張棟偵傛傝嵟傕憗婜偵僠儘僔儞儕儞巁壔偝傟傞儅僂僗抈敀傪惛惢扨棧偟SLP-65偲屇傫偩丅偝傜偵丄崟嶈抦攷愭惗傜偼撈帺偵僯儚僩儕BLNK傪扨棧偟丄BLNK寚懝DT40嵶朎傪嶌惢偟偰丄BLNK偑PLC兞2偺妶惈壔偵昁恵偱偁傞偙偲傪徹柧偟偨丅摨帪婜偵傂偲偮偺抈敀偑BASH, BLNK, SLP-65偲偄偆俁偮偺柤慜偱屇偽傟偰偄偨偙偲偵側傞丅Publication偺弴斣傪懜廳偟偰尰嵼偼BLNK偲屇傇丅
丂偙偺暘巕偺僲僢僋傾僂僩儅僂僗傕偦傟偧傟偺柦柤幰乮+ Kong-Peng Lam乯偺僌儖乕僾偱嶌惢偝傟偨偑丄枛徑儕儞僷宯偺昞尰宆偼丄惉弉B嵶朎傗暊峯B-1嵶朎偺寖尭丄BCR巋寖偵懳偡傞妶惈壔丒憹怋斀墳偺寚擛丄T嵶朎旕埶懚惈峈尨傊偺柶塽墳摎偺寚擛側偳丄Btk曄堎儅僂僗**偲崜帡偟偨丅偲偙傠偑丄崪悜偺昞尰宆偼Btk偁傞偄偼 preBCR寚娮儅僂僗偲傕堎側傝丄彫宆僾儗B嵶朎偑柍偔丄偦偺偐傢傝CD43偺敪尰偑傗傗掅壓偟丄偟偐傕昞柺preBCR梲惈偲偄偆偙傟傑偱偁傑傝尒偊側偐偭偨戝宆偺僾儗B嵶朎偑拁愊偟偰偄偨丅惓忢偺戝宆僾儗B嵶朎偼preBCR僔僌僫儖偵傛傝憹怋僒僀僋儖偵擖偭偰偄傞偑丄偙偺僾儗B嵶朎偼傎偲傫偳偑G0/G1婜偱掆巭偟偰偄偨丅偙偺寢壥偐傜丄BLNK偼僾儗B嵶朎偺暘楐偲偦偺屻偺彫宆壔傪桿摫偡傞preBCR僔僌僫儖揱払偵昁梫偱偁傞偲傢偐偭偨***丅
丂H嵔懳棫堚揱巕攔彍偵偼BLNK偼昁梫側偄偺偱丄Syk偵傛傞僔僌僫儖偺偡傋偰傪BLNK偑揱払偡傞傢偗偱偼側偄傛偆偩丅preBCR梲惈嵶朎偑拁愊偟偰偄傞偲偄偆偙偲偼preBCR偺down regulation偵傕BLNK傪夘偟偨preBCR僔僌僫儖偑摥偄偰偄傞偲峫偊傜傟傞丅傑偨丄彫宆僾儗B嵶朎偑柍偄偵傕偐偐傢傜偢怴惗B嵶朎偼寢峔偁傞偺偱丄L嵔堚揱巕嵞峔惉偼戝宆僾儗B嵶朎偱偐側傝偺昿搙偱婲偙偭偰偄傞偲巚傢傟傞丅偙傟偼RAG偺down regulation偑婲偙傜側偄偨傔偩傠偆偐丅偝傜偵丄帺屓峈尨偵斀墳偟偰receptor editing傪婲偙偟偰偄傞怴惗B嵶朎偼惓忢偱偼彫宆僾儗B嵶朎偲尒側偝傟偰偄傞偺偩偑丄彫宆僾儗B嵶朎偑柍偄偙偺儅僂僗偱偼偄偭偨偄receptor editing偼婲偙偭偰偄傞偺偩傠偆偐丅傕偟婲偙傜側偗傟偽丄偙偺儅僂僗偼帺屓柶塽昦偵側傞偺偩傠偆偐丅傛偆傗偔僐儘僯乕偑庽棫偱偒偨C57BL/6弮宯偺BLNK寚懝儅僂僗傪徻嵶偵夝愅偡傞偙偲偵傛偭偰丄崱屻偙傟傜偺媈栤傪夝寛偟偰偄偒偨偄丅傑偨丄Btk傕PLC兞2傕昁梫偱側偄偲偡傟偽BLNK偑偳偺傛偆偵preBCR僔僌僫儖傪壓棳偵揱偊偰偄傞偺偩傠偆偐丅偙傟偵偮偄偰傕丄巹払偑尒弌偟偨BLNK偺SH2僪儊僀儞偵寢崌偟偰妶惈壔偝傟傞HPK1偺婡擻傕娷傔偰丄BLNK傪夘偡傞僔僌僫儖揱払婡峔傪柧傜偐偵偟偨偄丅儕僈儞僪傪昁梫偲偟側偄偙偺帡旕庴梕懱偺僔僌僫儖揱払條幃偼偙傟傑偱偺忢幆偐傜偢偄傇傫奜傟偨傕偺側偺偐傕抦傟側偄丅
拲* 僸僩抝帣偵敪徢偡傞堚揱惈B嵶朎惈柶塽晄慡徢乮X-linked agamma-globulinemia: XLA, Bruton昦乯偱偼B嵶朎暘壔偑僾儗B嵶朎偱掆巭偟偰偄傞偑丄偦偺尨場偼Btk堚揱巕偺曄堎偱偁傞丅偟偨偑偭偰丄僸僩偱偼preBCR僔僌僫儖揱払偵Btk偑庡梫側摥偒傪偟偰偄傞偙偲偼柧傜偐偱偁傞丅
拲** 悢彮側偄柶塽宯偺帺慠曄堎儅僂僗偺偆偪丄惉弉B嵶朎偑彮側偔丄偦偺婡擻傕掅壓偟偰偄傞儅僂僗乮X-linked immunodeficiency: Xid乯偑抦傜傟偰偄偨偑丄偦偺尨場堚揱巕偑Btk偱偁偭偨丅屻偵嶌惢偝傟偨Btk堚揱巕僲僢僋傾僂僩儅僂僗傕傑偭偨偔摨偠昞尰宆偱偁偭偨丅
拲*** BLNK堚揱巕偵曄堎傪帩偮姵幰偱偼丄B嵶朎弶婜暘壔偑僾儗B嵶朎偱姰慡偵掆巭偟偰偄偨丅偟偨偑偭偰丄Btk偲摨條丄僸僩偵偍偄偰偼BLNK偼僾儗B嵶朎偺暘壔傪桿摫偡傞preBCR僔僌僫儖偵昁恵偱偁傞丅
乮摿掕椞堟尋媶乽柶塽僔僌僫儖揱払乿僯儏乕僗丄No.3, 暯惉12擭12寧宖嵹峞乯
乮堦晹夵曄乯
Back